Volume 27, Number 2, April 1998
Can Low Level Radiation Protect Against Cancer?
Ludwig E. Feinendegen, Victor P. Bond, Charles A. Sondhaus.
Abstract.Effects of ionizing radiation in living tissues are generally viewed in terms of damage. However, low doses of ionizing radiation also cause changes in normal intra- and extracellular signaling, with potentially beneficial consequences. Cellular signaling is triggered in part by reactive oxygen compounds and may result in the protection of cells against molecular damage, especially that to DNA. As shown in various rodent and human cell systems, radiation induced protective mechanisms serve the cell to defend against, repair, and remove damage to cells. Most of this damage comes from oxidative byproducts of normal metabolism. Only a relatively small fraction of the damage in tissues is caused by ionizing radiation, even at many times the dose from normal background radiation. Protective responses in various cell systems increase initially with dose up to about 0.1 - 0.2 Gy and then decrease and disappear. These responses are analyzed here in microdosimetric terms. The result contradicts in principle the linear nothreshold (LNT) hypothesis for radiation carcinogenesis. In fact, the incidence of spontaneous cancer may decrease at radiation doses below about 0.1 - 0.2 Gy. This is compatible with epidemiological data.
IntroductionThe risk RE of detrimental effects such as cancer induction from exposure of multicellular tissues to low doses of ionizing radiation is conventionally assumed to be proportional to the absorbed dose[21] D: R = alpha D (1) The expression in equation (1) formalizes the "linear nothreshold" (LNT) hypothesis. It states that even the smallest dose of radiation has a risk of causing cancer. The proportionality constant alpha has the dimension of inverse dose, Gy^-1 and is assumed to be of the order of 10^-4 Gy^-1 for cancer in human tissues exposed to low LET radiation. The average dose from background radiation at sea level is about 2 mGy per year. This paper analyses various effects of ionizing radiation in living tissues [15], and uses published data on low dose responses of tissues and cells in rodents and humans. The conclusions of this analysis contradict the LNT hypothesis. Living tissue is a nonlinear dynamic system, for which it would indeed be strange if such a simple expression of linearity were true over several orders of magnitude of dose.
Absorbed Dose to Tissue (Macrodose) and to Cells (Microdose)The absorbed dose D expresses energy/mass. In a macroscopic sense, the SI unit of dose is the gray, Gy, where 1 Gy = 1 J/kg. It has the convenience of being easily measured. However, at low values this macroscopic dose is inappropriate for estimating risk at the cellular level where cancer is induced. Thus, absorbed doses to individual cells, or microdoses, need to be considered. Ionizing radiation is absorbed in matter by the deposition of discrete amounts of energy along the tracks of charged particles that arise throughout the exposed matter [22]. The amount of energy absorbed from such tracks in a defined microvolume of tissue is here called a microdose. The microvolume of tissue is here taken to have the mass of a typical mammalian cell, 1 ng [3, 8, 12, 16, 28]. The mean microdose from a single interaction is calculated from the frequency of track events, or hits, in the defined micromass. Its value, z1, also called mean specific energy [22], hit-size or cell-dose, is a constant for a given type of radiation. For example, the z1 from 100 kV x-rays is about 1 mGy. According to microdosimetry theory [22], the tissue-absorbed dose D is equal to the product of z1 and N_H/N_e with N_H being the number of hits of any size, and N_e the number of exposed micromasses: D = z1 (N_H/N_e) (2) For the same macrodose from any type of radiation, N_H/N_e and z1 are reciprocally related. That is, for as given dose D of high LET radiation the average microdose z1 is proportionately greater and there are fewer hits for the same D than from low LET radiation. Substituting equation (2) into equation (1) gives RE = alpha . z1 . (N_H/N_e), and with RE being equal to the measured number of cancerous tumors N_q induced in the exposed tissue composed of _e cell-equivalent micromasses:[4] N_q/N_e= alpha z1 (N_H/N_e) This is known as the hit-number-effectiveness-function [4]. Multiplying each side of this equation by_ Ne yields: N_q = alpha . z1 . N_H. (3) Note that if the LNT hypothesis is correct, the proportionality constant alpha has the same value from equation 1 to 3. The Meaning of Dose RateIt has long been known that effects in irradiated living tissues depend on the rate at which the dose is absorbed. In microdose terms, the macrodose rate, Gy/unit time, allows one to determine how often on the average an individual cell is hit by a single average track event. The calculation [17] uses equation (2). Thus, D per unit time t is: D/t = z1 (N_H/N_e) 1/t or D/t = z1 /(t N_e/N_H) The denominator (t N_e/N_H) defines the average time interval between two consecutive hits per individual cell. This time interval also gives the average time available for repair of the cell prior to a second hit. This paper only considers short term exposures. A dose rate of 2 mGy per year from 100 kV x-rays would bring a hit to each individual cell in a 70 kg human on average once every 6 months. This may be viewed as two short term exposures of individual cells, 6 months apart. Since z1 from 100 kV x-rays is about 1 mGy, 2 mGy/y = 1 mGy/(y . 0.5)
Cellular Responses to MicrodosesThe response of living tissues to ionizing radiation in the low dose region involves various cellular responses [15] that are triggered mainly by hits in cells. Hit cells may also cause responses in non-hit cells by releasing signal substances and toxic, so-called clastogenic factors [9]. Moreover, hits in the extracellular matrix of the tissue may form products that interact with the cells [6] Indeed, the extracellular matrix may even reduce the natural incidence of cancer [2]. The response of tissues to low level radiation is, naturally, affected by tissue complexity. The quantification of cellular responses to hits in tissue is important for evaluating tissue risk at low doses. Cellular responses in tissue can be observed even after single hits per cell from low LET radiation [33]. These responses may have the following consequences [15]: a) Induction of damage: Radiation may change the structure of DNA and cause cancer. This is a major concern in radioprotection. High doses of ionizing radiation are known to increase the probability of cancer in the exposed tissues. The greater the dose, the larger is the risk. At low doses, less than 0.2 Gy of low LET radiation, no statistically significant increase of cancer is seen in mammalian tissues [35, 36]. 0.2 Gy corresponds to about 100 times the annual dose from background radiation at sea level and still causes about a 100 times less damage to the DNA than does normal cell metaboli[31]. The probability per average hit of inducing a cell in the exposed tissue to cause cancer is assigned the term p_ind. High-dose and high dose rate exposure exceeding about 0.3 Gy induces human leukemia proportionally to dose; at this high dose, p_ind is estimated to be approximately 10^-14 per average hit from low LET radiation in human blood forming stem cells [15]. Interestingly, p_ind per average hit in tissue culture cells [19] is about 10^-5 Thus, the difference between the two p_ind is a billion and is not readily explained only by the difference between the cellular mechanisms of malignant transformation in tissue and in culture. In this paper, p_ind is assumed to be constant over a wide range of doses from low LET radiation. If an enhancement of _ind were to result from an increase in the number of hits, then the enhanced probability of cancer per hit would then become p_ind (1+p_enh). For low LET radiation, p_enh per hit average is assumed to be zero. b) Activation of damage control: Low doses below about 0.2 Gy of low LET radiation affect physiological signaling within and between cells. This effect is due to chemical substances involving oxygen-containing radical compounds that are produced in the irradiated tissue. Altered signaling may change cellular metabolism in a variety of ways and trigger what can be viewed either as physiological responses of complex systems to toxic agents [10], or as adaptive responses [30]. They protect cells against damage to DNA and other biologically important molecules in the exposed tissue, irrespective of the cause of such damage. These protective responses may be grouped into four categories: 1) Damage prevention by temporarily stimulated detoxification of molecular radical species [11, 13, 14, 20]. This temporary protection of cellular constituents against toxic oxygen-containing radicals was found in mouse bone marrow at a maximum at 4 hours after short-term, i.e., acute, exposure to low doses of y-radiation; the degree of protection increased with dose up to about 0.1 Gy and then disappeared as a function of dose exceeding about -0.1 0.2[17,18]. 2) Damage repair by temporary stimulation of repair mechanisms. Low dose x-irradiation stimulated, i.e., conditioned the reduction of chromosomal aberrations that occur in cultures of human lymphocytes following large, so-called challenging doses [5, 30, 31, 33, 34, 38]. The degree of protection varied from zero to a maximum with individual donors of these cells. Protection was seen when the challenging dose of 2 Gy was given between about 4 and 70 hours after the conditioning dose of 0.005 - 0.01 Gy. It was not seen when the conditioning dose of 0.01 Gy was given at the very low dose rate of 0.0064 Gy/minute, or when the conditioning dose exceeded 0.1 Gy [32, 33], or when the challenging dose was 4 Gy instead of 2y [34]. 3) Damage removal by induction of apoptosis [23, 29]. Apoptosis is cell death in response mainly to DNA alterations; it is triggered by intracellular signals and eliminates damaged cells from tissue. At low doses of x-radiation, removal of damaged cells outweighed the induction of tissue damage from lost cells [23]. In one study, the incidence of this protective cell death in cultures of human lymphocytes rose up to day 4 after exposure to low LET radiation; it appeared linear with dose between about 0.1 and 2 Gy with a slope of 0.08 per 0.1 Gy [27]. 4) Damage removal by stimulated immune response [1, 26]. Cells of the immune system in rodents responded by stimulated production of Tcells during fractionated gamma-irradiation with 0.01 - 0.04 Gy per day for a total of 20 days [26]. The maximum response to acute whole-body gamma-irradiation was at doses between 0.1 - 0.3 Gy. This reponse improved surveillance of damaged cells over periods of weeks, and eliminated cancerous cells [1, 26]. An improved immune response may result in an increased resistance to common infections and prolong life [37]. The four protective responses all activate cell damage control and are here summed up by a cumulative probability p_prot per average hit. Damage to cells results mainly from oxidative byproducts of normal metabolism and not from low dose irradiation[31]. Thus, radiation-induced protection acts mainly by preventing such damage. Also, existing damage may be eliminated by protective responses. Because protection may be mediated by way of intercellular signaling and by the extracellular matrix, nonirradiated cells may also benefit. Since DNA damage may induce cancer [7, 37], the radiation-induced protective responses may also protect against spontaneously occurring cancer [37]. The probability of a spontaneous malignant transformation per cell is denoted here by p_spo. In the human blood forming stem cells, about 10^9 in the adult, p_spo per cell causing leukemia is approx^-11 throughout life [17]. As stated above for the human blood forming stem cell, p_ind per average hit is about 10^-14. Hence, the ratio p_spo/p_ind is about 10^3. This is a factor of ten smaller than the ratio of metabolically produced DNA damage and that due to background radiation per cell per day [31]. Regardless of the mechanisms involved, the values of p_spo, p_ind and p_prot are likely to vary with the organism, cell type, and z 1. This emphasizes the principal difficulty of predicting the risk of cancer for an individual from a given dose of radiation. Nevertheless, in contrast to _ind, in different cell systems p_prot decreased when D exceeded about 0.1 to 0.2 Gy of low LET radiation, as was discussed above [17,32,33,34]. The observed detrimental and protective responses in the exposed tissue operate transiently over different time spans. Thus, in order to compare the various p-values numerically, their averages need to be established and normalized to their time of duration. Moreover, a radiation-induced malignant transformation in a single cell may eventually cause a tumor to develop only over a period of perhaps several decades and following a series of subsequent DNA mutations. Therefore, in order to offset one randomly occurring spontaneous transformation leading to later tumor development, either the respective cell must experience a temporary protective response often or an accordingly large number of respective cells must each be temporarily protected once.
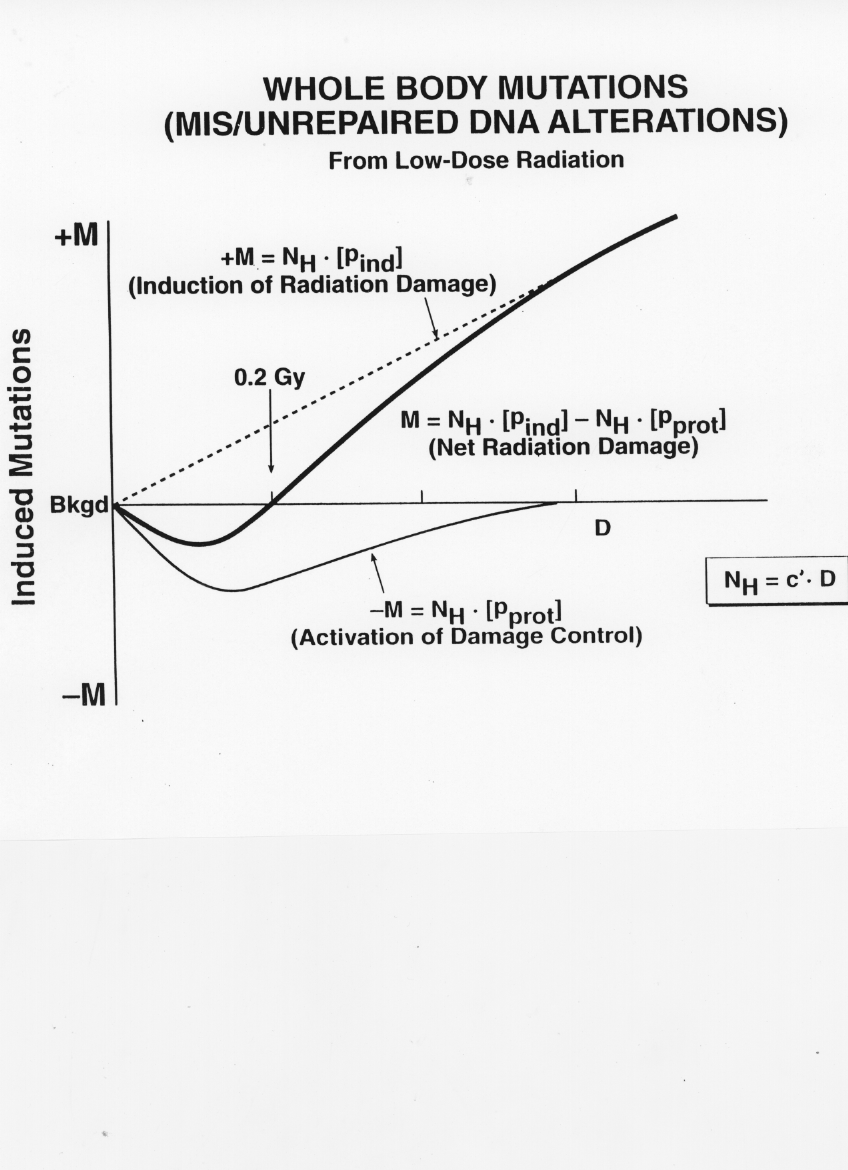
Tissue Response as the Sum of Microdose EffectsThe preceding section indicates that low doses of low LET radiation that result in single or few hits per cell may initiate cancer and also produce various protective effects against both spontaneous and radiation induced cancer. The following probabilities per cell are here defined: p_spo = spontaneous malignant transformation throughout life, p_ind = radiation induced malignant transformation per average hit, p_enh = enhancement of p_ind per average hit, p_prot = protection that will prevent cancer from developing in the exposed tissue, per average hit. The total probability per average hit of causing a cancer in the exposed tissue can then be expressed as [18]: N_q/N_H = [p_ind + p_ind p_enh - p_prot p_spo - p_prot p_ind- p_prot p_ind p_enh] (4) With N_q/N_H = alpha . z1 (see equation 3) and rearranging equation (4) : alpha = [p_ind (1 + p_enh)- p_prot(p_spo+p_ind+p_ind p_enh)](1/z1) (5) In Figure 1, equation (5) is abbreviated to alpha = [{P_ind} - {P_prot}] (1/z1) (6) with { P_ind} = p_ind (1 + p_enh) ; and {P_prot} = p_prot(p_spo+p_ind+p_ind p_enh). For low LET radiation, p_enh is taken to be zero. {P_prot} increases up to a dose of about 0.1 - 0.2 Gy. At higher doses it decreases gradually to zero. Since the protective term goes through a maximum in the low dose range, it is impossible for alpha to be a constant as required for the LNT hypothesis [15, 17, 1]. The various p-values estimated above may be used for a crude approximation of the probability of radiation induced leukemia. Thus, the value of p_ind for blood forming stem cells is about 10^-14 per hit average of low LET radiation [15]; p_enh is considered zero for low-LET radiation; p_spo for the same cell type has been estimated to be about 10^-11 throughout life [17]; thus, the value of the negative term in alpha, p_prot (p_spo + p_ind +p_ind p_enh) would become equal to the value of the positive term p_ind (1 + p_enh) if p_prot were 10^-3 at some dose of low-LET radiation. At this dose, there would be no increase in leukemia and a threshold would exist. With higher values of p_prot the leukemia incidence would be lower than in the control population; conversely, RE in equation (1) would only increase with D, as p_prot decreases following a maximum (see Fig. 1). Thus, with regard to radiation-induced cancer, at least two different dose effect curves must be considered. According to equation (6) and as seen in Figure 1, the sum of these dose effect curves generates the net dose effect curve. In Figure 1, D is the product of N_H and the constant z1/N_e making N_H = c' . D. On the vertical axis is the increase (+M) or decrease (-M) of cancers in the exposed tissue compared to the background cancer incidence. Most of these background cancers are due to DNA damage resulting from normal cellular metabolism. Components of p_prot are easily measured at low doses in various cell systems whereas _ind is not [37]. If cancer is induced at low doses, i.e., below about 0.2 Gy, it is lost in the statistical noise of spontaneous cancer incidence. Indeed, a lack of statistically significant changes in N_q after exposure of mammalian tissues to low LET radiation below 0.2 Gy makes it impossible to determine whether detrimental, beneficial, or no effects occur [36, 37]. However, epidemiological and many experimental data support the existence of a threshold or even beneficial (hormetic) effects at low D and low LET radiat[24,25]. Regarding the term alpha for high LET radiation, the relatively high value of z1 may cause p_prot for the exposed cells to be so small as to be ineffective. However, p_ind, p_ind p_enh and p_spo might be offset by a larger p_prot if it is initiated in non-hit cells by intercellular signal substances and specific toxic factors stemming from irradiated cells. Stimuli that are induced by high LET radiation in the extracellular matrix might also affect nonhit cells.
ConclusionFor assessing the probability of radiation induced cancer at low doses, the absorbed dose of importance is that to individual micromasses, the microdose. The macrodose, D, has limited applicability in estimating cancer risk at low doses. The macrodose from a given type of ionizing radiation in tissue is equal to the product of the number of microdose events, or hits, N_H, per number of exposed micromasses, N_e and the average energy absorbed per hit micromass, z1, i.e., the mean microdose for that radiation. For the calculation of z1 for any type of radiation, the micromass in tissue is taken to be 1 ng and corresponds to the mass of a typical cell in mammalian tissue. For the same macrodose from any type of radiation, N_H/N_e and z1 are reciprocally related. That is, for a dose D for high LET radiation, the average microdose z1 is proportionately greater with fewer hits than for the same D of low LET radiation. The structure and function of tissues are determined by cells, the elemental units of life. Cells respond to hits. Adjoining cells may be affected by signal substances from hit cells and irradiated extracellular matrix. One type of response to low LET irradiation initiates cellular damage, especially to DNA. Other responses protect against or remove DNA damage by way of stimulating existing cellular mechanisms for DNA damage control. The mis- or unrepaired DNA damage that is constantly caused by oxidative by-products of normal metabolism is estimated to be about 10,000 times greater than DNA damage from background radiation [31] Mis- or unrepaired DNA damage may cause cancer. It follows that the protection produced by hit cells mainly operates against spontaneous cancer from normal metabolism. Low dose of low LET irradiation of different mammalian cell systems shows that the protective mechanisms increase initially with dose up to about 0.1 Gy. As the dose increases above 0.1 0.2 Gy, the protective action decreases and eventually disappears. The biphasic response of the protective action of radiation is in marked contrast to the assumed linear increase of cancer with low doses predicted by the LNT hypothesis. The incidence of radiation induced cancer may be zero at low doses and the spontaneous cancer incidence may even fall in the dose region around about 0.1 Gy. Many epidemiological and experimental data support this conclusion.
References1. Anderson R.E. Effects of low-dose radiation on the immune response in: Biological effects of low level exposures to chemicals and radiation. Ed. E.J. Calabrese. Lewis Pub. Inc. Chelsea, Michigan. 1992, p.95-112 2. Bissel M.J. personal communication, 1996 3. Bond V.P., Feinendegen L.E. Intranuclear 3-H thymidine: Dosimetric, radiobiological and radioprotection aspects. Health Phys. 12: 1007-1020 (1966) 4. Bond V.P., Benary V., Sondhaus C.A., Feinendegen L.E. The meaning of linear dose-response relations, made evident by use of absorbed dose to the cell. Health Phys. 68: 8B6-792 (1995) 5. Bosi A., Olivieri G. Variability of the adaptive response to ionizing radiations in humans. Mutation Res. 211: 13-17 (1989). 6. Boudreau, N., Werb Z., Bissel M.J. Suppression of apoptosis by basement membrane requires threedimensional tissue organization and withdrawal from the cell cycle. Proc.Natl.Acad.Sci.USA 93: 3509-3513 (1996) 7. Cleaver J. Defective repair replication of DNA in Xeroderma Pigmentosum. Nature (London) 21B: 652-656 (1968). B. Cronkite E.P., Robertson J.S., Feinendegen L.E. Somatic and teratogenic effects of tritium. in: Tritium. Moghissi A.A., Carter M.W. eds. Messenger Graphics, Phoenix AZ, 198-209 (1971) 9. Emerit I., Oganesian N., Sarkisian T., Arutyunyan R., Pogosian A., Asrian K., Levy A., Cernjavski L. Clastogenic factors in the plasma of Chernobyl accident recovery workers: Anticlastogenic effect of Gingko Biloba extract. Radiation Res. 144: 19B-205 (1995) 10. Feinendegen L.E., Muehlensiepen H., Lindberg C., Marx J., Porschen W., Booz J. Acute effect of very low dose in mouse bone marrow cells: a physiological response to background radiation? in: Biological Effects of Low Level Radiation, IAEA Vienna 459- 471 (1983). 11. Feinendegen L.E., Muehlensiepen H., Lindberg C., Marx J., Porschen W., Booz J. Acute and temporary inhibition of thymidine kinase in mouse bone marrow cells after low-dose exposure. Int.J.Radiat.Biol. 45: 205-215 (1984). 12. Feinendegen L.E., Booz J., Bond V.P., Sondhaus C.A. Microdosimetric approach to the analysis of cell responses at low dose and low dose rate. Radiat.Protect.Dosimetry 13: 299-306 (1985). 13. Feinendegen L.E., Muehlensiepen H., Bond V.P., Sondhaus C.A. Intracellular stimulation of biochemical control mechanisms by low-dose low-LET irradiation. Health Phys. 52: 663-669 (1987). 14. Feinendegen L.E., Bond V.P., Booz J., Muehlensiepen H. Biochemical and cellular mechanisms of low-dose effects. Int.J.Radiat.Biol. 53: 23-37 (1988). 15. Feinendegen L.E. Radiation risk of tissue late effect, a net consequence of probabilities of various cellular responses. Europ.J.Nucl.Med. 1B: 740-751 (1991) 16. Feinendegen L.E., Bond V.P., Booz J. The quantification of physical events within tissue at low levels of exposure to ionizing radiation. ICRU-News 2: 9-13 (1994) 17. Feinendegen L.E., Loken M., Booz J., Muehlensiepen H., Sondhaus C.A., Bond V.P. Cellular mechanisms of protection and repair induced by radiation exposure and their consequences for cell system responses. Stem Cells 13(1): 7-20 (1995) 1B. Feinendegen L.E., Bond, V.P., Sondhaus C.A., Muehlensiepen H. Radiation effects induced by low doses in complex tissue and their relation to cellular adaptive responses. Mutation Res. 35B: 199-205 (1996) 19. Hall E.J., Hei T.K. Oncogenic transformation in vitro by radiation of varying LET. Radiat.Protect.Dosimetry 13: 149-151 (1985). 20. Hohn-El-Karim K., Muehlensiepen H., Altman K.I., Feinendegen L.E. Modification of effects of radiation on thymidine kinase. Intern.J.Radiat.Biol. SB: 97-110 (1990) 21. ICRU (International Commission on Radiation Units and Measurements) Radiation Quantities and Units. ICRU, Bethesda, MD, Report 33 (1980). 22. ICRU (International Commission on Radiation Units and Measurements) Microdosimetry. ICRU, Bethesda, MD, Report 36 (1983). 23. Kondo S. Health Effects of Low Level Radiation. Kinki Univ.Press, Osaka, Japan; Medical Physics Publishing, Madison, WI; 1993. 24. Loken M.K., Feinendegen, L.E. Radiation hormesis, its emerging significance in medical practice. Invest.Radiol. 2B: 446-450 (1993) 25. Luckey T.D. Physiological benefits from low level of radiation exposure. Health Phys. 43: 771-7B5 (1982). 26. Makinodan T. Cellular and subcellular alteration in immune cells induced by chronic, intermittent exposure in vivo to very low dose of ionizing radiation (ldr) and its ameliorating effects on progression of autoimmune disease and mammary tumor growth. In: Low-Dose Irradiation and Biological Defense Mechanisms; Amsterdam; London; New York; Tokyo. Exerpta Medica 1992, 233-237 27. Menz R.C. An assay for biological dosimetry based on induction of apoptosis in human T-lymphocytes. Thesis, St. Bartholomew's and the Royal London School of Medicine and Dentistry, University of London, 1996. 28. NCRP (National Council on Radiation Protection) . Tritium and Other Radionuclide Labeled Organic Compounds Incorporated in Genetic Materials. NCRP, Bethesda MD, Publication 63 (1979) 29. Norimura T., Nomoto S., Katsuki M., Gondo Y., Kondo S. p-5 3-dependent apoptosis suppresses radiation-induced teratogenesis. Nat.Med. 2: 577-580 (1996) 30. Olivieri G., Bodycote J., Wolff S. Adaptive response of human lymphocytes to low concentrations of radioactive thymidine. Science 223: 594-597 (1984) 31. Pollycove M., Feinendegen, L.E. Quantification of human total and mis/unrepaired DNA alterations: intrinsic and radiation induced. To be published 32. Shadley J.D., Wolff S. Very low doses of X-rays can induce human lymphocytes to become less susceptible to ionizing radiation. Mutagenesis 2: 95-96 (1987). 33. Shadley J.D., Afzal V., Wolff S. Characterization of the adaptive response to ionizing radiation induced by low doses of X-rays to human lymphocytes. Radiation Res. 111: 511-517 (1987) 34. Shadley J.D., Wienke, J.K. Induction of the adaptive response by X-rays is dependent on radiation intensity. Int.J.Radiat.Biol. 56: 107-liB (1989). 35. Shadley J.D., Dai G. Cytogenetic and survival adaptive responses in G phase human lymphocytes. Mutation Res. 265: 273-2B1 (1992) 36. UNSCEAR. Ionizing Radiation: Sources and Biological Effects. United Nations, New York, N.Y.: 19BB. 37. UNSCEAR. Sources and Effect of Ionizing Radiation. United Nations, New York, N.Y.: 1994 38. Wei Q., Matanoski G.M., Farmer E.R., Hedayati M.A., Grossman L. DNA repair and aging in basal cell carcinoma: A molecular epidemiologic study. Proc.Natl.Acad.Sci.USA 90: 1614-161B (1993) 39. Wolff S., Afzal V., Wienke J.K., Olivieri G., Michaeli A. Human lymphocytes exposed to low doses of ionizing radiations become refractory to high doses of radiation as well as to chemical mutagens that induce double-strand breaks in DNA. Int.J.Radiat.Biol. 53(1): 39-49 (1988). 40. Zamboglou N., Porschen W., Muehlensiepen H., Booz J., Feinendegen L.E. Low dose effect of ionizing radiation on incorporation of iododeoxyuridine into bone marrow cells. Int.J.Radiat.Biol. 39: 83-93 (1981). |
{kind=link}